 Problem 4: Naturlig utvalg sliter med å fiksere fordelaktige egenskaper i populasjoner
Problem 4: Naturlig utvalg sliter med å fiksere fordelaktige egenskaper i populasjoner
Bilde 1. Enda nyere konferanse (2016)
I 2008 ble 16 biologer fra hele verden samlet i Altenberg, Østerrike for å diskutere problemer med den moderne neo-darwinistiske evolusjonsmodellen. Tidsskriftet Nature dekker denne "Altenberg 16" konferansen, og siterer ledende forskere som sier ting som:
"Opprinnelsen til vinger og invadering av landjorda. . . er ting som evolusjonsteorien har fortalt oss lite om." (49)
"Du kan ikke nekte kraften til seleksjon i genetisk evolusjon. . . men etter min mening er dette stabiliserende og finjusterende former som kommer fra andre prosesser. "
 "Den moderne syntesen er bemerkelsesverdig god til å modellere overlevelsen til de mest tilpassede, men ikke god til å modellere ankomsten til de mest tilpassede."
"Den moderne syntesen er bemerkelsesverdig god til å modellere overlevelsen til de mest tilpassede, men ikke god til å modellere ankomsten til de mest tilpassede."
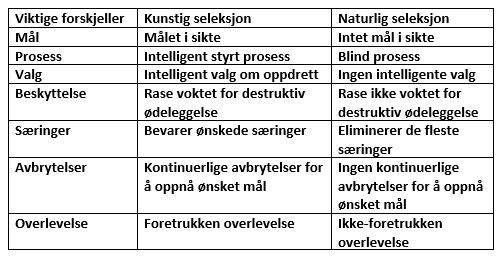
I problem 3 -her; lærte vi at mutasjoner ikke kan generere mange komplekse egenskaper i levende organismer i løpet av rimelige evolusjonære tidsskalaer. Men mutasjoner er bare en del av standard evolusjonær mekanisme - det finnes også naturlig utvalg. Og den darwinistiske evolusjonen forklarer ikke bare ofte 'den best tilpassede' ankomst via mutasjoner, men sliter også ofte med å forklare 'overlevelse til de mest tilpassede' via naturlig utvalg.
Bilde 3. Opphavsmann til 'naturlig seleksjon'
 Evolusjonære biologer antar ofte at når mutasjoner produserer en funksjonelt fordelaktig egenskap, vil den lett spre seg (bli 'fiksert') gjennom en populasjon via naturlig utvalg. For eksempel, tenk på en befolkning av brunhåret rev som bor i en snø-region. En rev er født med en mutasjon som gjør at pelsen er hvit, i stedet for brun. Denne reven har nå en fordel i å jakte bytte og unngå rovdyr, fordi den hvite pelsen gir den kamuflasje i det snøfylte miljøet. Den hvite rev overlever, avleverer sine gener til sitt avkom, som også er dyktige i å overleve og reprodusere. Over tid spres den hvithårede egenskapen i hele befolkningen.
Evolusjonære biologer antar ofte at når mutasjoner produserer en funksjonelt fordelaktig egenskap, vil den lett spre seg (bli 'fiksert') gjennom en populasjon via naturlig utvalg. For eksempel, tenk på en befolkning av brunhåret rev som bor i en snø-region. En rev er født med en mutasjon som gjør at pelsen er hvit, i stedet for brun. Denne reven har nå en fordel i å jakte bytte og unngå rovdyr, fordi den hvite pelsen gir den kamuflasje i det snøfylte miljøet. Den hvite rev overlever, avleverer sine gener til sitt avkom, som også er dyktige i å overleve og reprodusere. Over tid spres den hvithårede egenskapen i hele befolkningen.
Slik fungerer det - i teorien. Men i den virkelige verden garanterer ikke bare å generere en funksjonelt fordelaktig egenskap, at den vil vedvare eller bli fiksert.
For eksempel, om ved en tilfeldighet den hvite reven, brekker et bein og blir spist av en rovdyr - aldri passerer sine gener? Tilfeldige krefter eller hendelser kan forhindre at et trekk sprer seg gjennom en populasjon, selv om de gir en fordel. Disse tilfeldige kreftene grupperes sammen under navnet "genetisk drift". Når biologer anvender matematikk på naturlig utvalg, finner de at med mindre et trekk gir en ekstremt sterk selektiv fordel, vil genetisk drift ha en tendens til å overvelde styrken ved seleksjon og hindre tilpasninger fra å få fotfeste i en populasjon.
Dette undervurderte problemet er blitt erkjent av noen evolusjonære forskere, som er skeptiske til evnen av naturlig utvalg for å drive den evolusjonære prosessen. En av disse forskerne er Michael Lynch, en evolusjonær biolog ved Indiana University, som skriver at "tilfeldig genetisk drift kan fremsette en sterk barriere for fremdriften til molekylære forbedringer ved hjelp av adaptive prosesser." (50) Han bemerker at virkningen av seleksjon er "oppmuntrende til fiksering av mildt skadelige mutasjoner og motvirker fremme av gunstige mutasjoner. " (51) Eugene Koonin, en ledende forsker ved National Institutes of Health, forklarer også at genetisk drift fører til 'tilfeldig fiksering av nøytrale eller skadelige endringer'. (52)
Kompleks redundans
 I Lynchs syn er det mange cellulære systemer som bidrar til overlevelse, men er overflødige. Som et resultat tjener de som backup mekanismer som bare brukes når et svært effektivt primærsystem mislykkes. Fordi de bare er sjelden brukt, blir disse systemene bare av og til eksponert for siktingen ved seleksjon. Likevel kan disse systemene være svært komplekse og effektive. Hvordan kan et system som bare sjelden brukes, eller bare nødvendigvis, utvikle seg til et så høyt og effektivt nivå av kompleksitet? Etter å ha observert de mange 'lagene' av komplekse cellulære mekanismer som er involvert i prosesser som DNA-replikasjon, stiller Lynch et avgjørende spørsmål:
I Lynchs syn er det mange cellulære systemer som bidrar til overlevelse, men er overflødige. Som et resultat tjener de som backup mekanismer som bare brukes når et svært effektivt primærsystem mislykkes. Fordi de bare er sjelden brukt, blir disse systemene bare av og til eksponert for siktingen ved seleksjon. Likevel kan disse systemene være svært komplekse og effektive. Hvordan kan et system som bare sjelden brukes, eller bare nødvendigvis, utvikle seg til et så høyt og effektivt nivå av kompleksitet? Etter å ha observert de mange 'lagene' av komplekse cellulære mekanismer som er involvert i prosesser som DNA-replikasjon, stiller Lynch et avgjørende spørsmål:
-Selv om disse lagdelte forsvarslinjene er klart fordelaktige og i mange tilfeller avgjørende for cellehelsen, fordi samtidig oppkomst av alle komponentene i et system er usannsynlige, oppstår flere spørsmål umiddelbart. Hvordan kan seleksjon fremme etablering av flere lag med tilpasnings-fremmende mekanismer hvis de etablerte primære forsvarslinjene allerede er svært raffinert? (53)
Lynch tror ikke naturlig utvalg mesterer oppgaven. I et 2007-dokument i Proceedings of the U.S. National Academy of Sciences med tittelen " Svakheten ved adaptive hypoteser for opprinnelsen til organisme-kompleksitet", forklarer han at blant evolusjonære biologer: "Det som er spørsmålet er om naturlig utvalg er en nødvendig eller tilstrekkelig kraft for å forklare fremveksten av genomiske og cellulære funksjoner som er sentrale for bygging av komplekse organismer." (54) Ved bruk av lignende språk, konkluderer en artikkel i tidsskriftet Theoretical Biology and Medical Modelling at "det er viktig for biologer å realistisk vurdere hva naturlig utvalg kan og ikke kan gjøre under ulike omstendigheter. Seleksjon kan vise seg ikke å være nødvendig eller tilstrekkelig til å forklare mange genomiske eller cellulære trekk ved komplekse organismer." (55) Lynch er tydelig i  sine synspunkter:" Det er ingen tvingende empiriske eller teoretiske bevis på at kompleksitet, modularitet, redundans eller andre egenskaper av genetiske stier fremmes ved naturlig utvalg." (56)
sine synspunkter:" Det er ingen tvingende empiriske eller teoretiske bevis på at kompleksitet, modularitet, redundans eller andre egenskaper av genetiske stier fremmes ved naturlig utvalg." (56)
Enten du appellerer til seleksjon eller ikke: du vinner ikke.
I stedet for naturlig utvalg, foreslår evolusjonære biologer som Lynch tilfeldig genetisk drift til å forklare opprinnelsen til komplekse biologiske egenskaper. Ifølge Lynch er "mange aspekter av kompleksitet på genomiske, molekylære og cellulære nivåer i multi-cellulære arter skylder trolig til sin opprinnelse til disse ikke-adaptive kreftene, noe som representerer lite mer enn passive resultater ..." (57) Men han anerkjenner at disse 'ikke-adaptive krefter' av evolusjon er stokastiske av natur." (58)
Stokastisk betyr selvfølgelig tilfeldig. Kan en strengt tilfeldig kraft - som ikke har grunn til å bevare egenskaper som kan gi noen fordel - forklare de svært komplekse biologiske egenskapene - som DNA-replikasjon eller bioluminescens - som synes finjustert til å utføre nyttige biologiske funksjoner? Biolog Ann Gauger er skeptisk til Lynchs forklaring, da hun observerer at han "gir ingen forklaring på hvordan ikke-adaptive krefter kan produsere den funksjonelle genomiske og organisatoriske kompleksiteten vi observerer i moderne arter." (59) Jerry Coyne påpeker på samme måte den store mangelen i appeller til genetisk drift:
-Både drift og naturlig utvalg gir genetisk forandring som vi anerkjenner som evolusjon. Men det er en viktig forskjell. Drift er en tilfeldig prosess, mens seleksjon er anti-tesen av tilfeldighet. ... Som en ren tilfeldig prosess kan genetisk drift ikke føre til utviklingen av tilpasninger. Det kunne aldri bygge en vinge eller et øye. Det krever ikke-tilfeldig naturlig seleksjon. Det drift kan gjøre er å utvikle trekk som verken er nyttige eller skadelige for organismen. (60)
 Coyne observerer videre : "Innflytelsen av denne prosessen på viktig evolusjonær endring er imidlertid sannsynligvis liten, fordi den ikke har formkraften til naturlig utvalg. Naturlig utvelgelse er fortsatt den eneste prosessen som kan gi tilpasning." (61) Men på en måte enig med Lynch, innser han at "genetisk drift ikke bare er maktesløs i å skape tilpasninger, men faktisk kan overvinne naturlig utvalg." (62)
Coyne observerer videre : "Innflytelsen av denne prosessen på viktig evolusjonær endring er imidlertid sannsynligvis liten, fordi den ikke har formkraften til naturlig utvalg. Naturlig utvelgelse er fortsatt den eneste prosessen som kan gi tilpasning." (61) Men på en måte enig med Lynch, innser han at "genetisk drift ikke bare er maktesløs i å skape tilpasninger, men faktisk kan overvinne naturlig utvalg." (62)
Debatten om hvorvidt naturlig utvalg , eller genetisk drift, er mest innflytelsesrik i evolusjonen, fortsetter utvilsomt. Men det er liten grunn til å tro at hvilken side som enn vinner denne debatten, vil det bli tilbudt en levedyktig materialistisk løsning. Den evolusjonære biologien befinner seg nå overfor en catch-22:
Naturlig utvalg er for ineffektiv en mekanisme i å overvinne tilfeldige krefter og fiksere de komplekse tilpasninger vi observerer i populasjoner, fordi den lett overstyres av tilfeldige krefter som genetisk drift.
Livet er fullt av svært komplekse og effektive tilpasninger, men tilfeldig genetisk drift gir ingen forsvarlig grunn til å tro at slike funksjoner vil ha noen grunn til å oppstå.
I hovedsak er genetisk drift lik å påkalle "mutasjon-seleksjons" -mekanismen, men minus alt av seleksjon. Disse emnene peker på alle de vanskelighetene vi så i problem 3 -lenke, hvor tilfeldige mutasjoner ikke klarte å bygge biokjemiske egenskaper som funksjonelle proteiner eller enkle protein-protein-interaksjoner, fordi det var nødvendig med flere koordinerte mutasjoner for å produsere disse egenskapene. Mangler man seleksjon, er det ingen grunn for tilfeldige mutasjoner alene - dvs. genetisk drift - til å produsere noe som er nyttig.
 Dessverre blir publikum sjelden kjent med disse problemene eller denne debatten. Ifølge Lynch, er naturlig utvalg vanligvis portrettert som en 'allmektig kraft (uten direkte bevis)' (63) mekanisme som kan bygge komplekse biologiske egenskaper. Han advarer om at "myten om at all evolusjon kan forklares ved tilpasning fortsetter å bli videreført ved vår fortsatte respekt, via Darwins behandling i populærlitteraturen." (64) Virkeligheten er at verken ikke-tilfeldige krefter som naturlig utvalg eller tilfeldige krefter som genetisk drift, kan forklare opprinnelsen til mange komplekse biologiske egenskaper.
Dessverre blir publikum sjelden kjent med disse problemene eller denne debatten. Ifølge Lynch, er naturlig utvalg vanligvis portrettert som en 'allmektig kraft (uten direkte bevis)' (63) mekanisme som kan bygge komplekse biologiske egenskaper. Han advarer om at "myten om at all evolusjon kan forklares ved tilpasning fortsetter å bli videreført ved vår fortsatte respekt, via Darwins behandling i populærlitteraturen." (64) Virkeligheten er at verken ikke-tilfeldige krefter som naturlig utvalg eller tilfeldige krefter som genetisk drift, kan forklare opprinnelsen til mange komplekse biologiske egenskaper.
Referanser:
[49.] Scott Gilbert, Stuart Newman, and Graham Budd quoted in John Whitfield, “Biological theory: Postmodern evolution?” Nature, 455: 281-284 (September 17, 2008).[50.] Michael Lynch, “Evolutionary layering and the limits to cellular perfection,” Proceedings of the U.S. National Academy of Sciences, www.pnas.org/cgi/doi/10.1073/pnas.1216130109(2012).
[51.] Michael Lynch, “The frailty of adaptive hypotheses for the origins of organismal complexity,” Proceedings of the U.S. National Academy of Sciences, 104: 8597-8604 (May 15, 2007).
[52.] Eugene V. Koonin, “Darwinian evolution in the light of genomics,” Nucleic Acids Research(2009): 1-24, doi:10.1093/nar/gkp089 [53.] Ibid.
[54.] Michael Lynch, “The frailty of adaptive hypotheses for the origins of organismal complexity,” Proceedings of the U.S. National Academy of Sciences, 104: 8597-8604 (May 15, 2007).
[55.] Chase W. Nelson and John C. Sanford, “The effects of low-impact mutations in digital organisms,” Theoretical Biology and Medical Modelling, 8:9 (2011).
[56.] Michael Lynch, “The evolution of genetic networks by non-adaptive processes,” Nature Reviews Genetics, 8:803-813 (October, 2007). [57.] Ibid.
[58.] Michael Lynch, “The frailty of adaptive hypotheses for the origins of organismal complexity,” Proceedings of the U.S. National Academy of Sciences, 104: 8597-8604 (May 15, 2007).
[60.] Jerry A. Coyne, Why Evolution is True, p. 123 (Viking, 2009). [61.] Ibid., p. 13. [62.] Ibid., p. 124.
[63.] Michael Lynch, “The frailty of adaptive hypotheses for the origins of organismal complexity,” Proceedings of the U.S. National Academy of Sciences, 104: 8597-8604 (May 15, 2007). [64.] Ibid.
Oversettelse og bilder ved Asbjørn E. Lund